Advanced Epigenetic Overview of Histone Modifications
A detailed guide to histone modifications investigated in epigenetic research
Histone modifications involve covalent post-translational modifications (PTMs) mostly to the residues at the positively charged N-terminal tails of histones. The most common histone modifications are methylation, acetylation and phosphorylation. Some other modifications include sumoylation, ribosylation, ubiquitylation, and citrullination among others.
Histone modifications regulate gene expression by changes to the chromatin structuredue to the disruption of the electrical charges in the histone residues, relaxing or compacting chromatin; and by recruiting DNA binding proteins, with subsequent silencing or activating effects. Looser, more relaxed chromatin, known as euchromatin, is more accessible for transcription, resulting in increased gene expression. Whereas heterochromatin, on the other hand, is more tightly compacted, making it less accessible for transcription leading to reduced gene expression. Besides their direct impact on the chromatin structure, they also interact and affect the establishment of other epigenetic marks. For instance, specific histone modifications are key for establishing DNA methylation patterns.
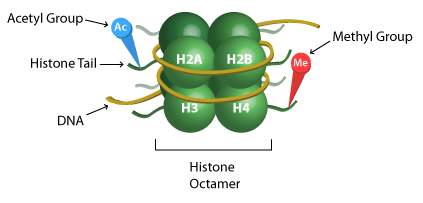
Histones H3 and H4 undergo the majority of these modifications, although histones H2A and H2B also are subject to some alterations. The enzymes that catalyze these changes are diverse, and include histone methyltransferases (HMTs), and histone acetylases (HATs) among others. These changes, on the other hand, are reversible and specific enzymes catalyze their removal. For instance, histone demethylases (HDMTs) and histone deacetylases (HDACs) catalyze the removal of the methylation and acetylation marks in histones, respectively. Besides post-translational modifications at the histones N-terminal tails, the core domains of the histones also undergo modifications.
Histones are acetylated by addition of an acetyl group to the Ɛ-amino group on lysine residues by histone acetyltransferases (HATs). Histone acetylation is associated with transcriptional activation and is thus an activation mark, as acetylation neutralizes the positive charge of lysine residues and causes the weakening of DNA and histone interactions leading to “looser” chromatin. On the other hand, histone deacetylases (HDACs)catalyze the removal of the acetyl groups from the histones, and lead to transcriptional repression.
Histones are methylated by transfer of one to three methyl groups to the side chains of lysine or arginine residues by histone methyltransferases (HMTs). Lysines can be mono, di, or tri-methylated, whereas arginines can be mono or di-methylated. Unlike acetylation and phosphorylation, histone methylation does not change the positive charge of the amino acid residues. These methyl groups can act as activating or repressive marks. For example, the tri-methylation of histone H3 at lysine 27 (H3K27me3) leads to silencing and repression of gene expression; whereas the tri-methylation of histone H3 at lysine 4 (H3K4me3) leads to transcriptional activation. On the other hand, histone demethylases (HDMTs) catalyze the removal of the methylation marks. For lysine, lysine specific demethylases (LSDs) and JmJC-domain containing proteins are the 2 groups of enzymes responsible for reversal of the mark. For arginine, it is thought that a few JmJC-domain containing enzymes catalyze the reversal.
Histone Phosphorylation
Histones can also be phosphorylated at serine, threonine, and tyrosine residues by kinases. Phosphorylation imparts a highly negative charge to the residues and affects the chromatin structure by generally promoting a looser chromatin. Important phosphorylation modifications include phosphorylation of histone H2AX (γH2AX), an important marker of DNA damage. Several kinases mediate the phosphorylation of histones, among them Aurora B, while certain phosphatases, like phosphatase PP1, dephosphorylate and revert this modification.
Other histone PTMs, include ribosylation, ubiquitination, sumoylation, citrullination and glycosylation, among others. ADP ribosylation consists of the addition of one or more ADP-ribose on glutamate and arginine residues. It imparts a negative charge and is associated with a looser chromatin structure. Mono ADP-ribosylation is catalyzed by the mono-ADP-ribosyltransferases, whereas poly-ADP-ribosylation is catalyzed by the PARP family of enzymes.
Ubiquitination results in a much larger modification, as ubiquitin is 76 amino-acids long. Lysine residues can be mono or poly-ubiquitinated. Histones are mostly monoubiquitinated, in particular Histones H2A and H2B, and these changes can be repressive or activating.
Sumoylation is a modification similar to ubiquitination and consists of attachment of small ubitiquin-like modifier (SUMO) molecules to lysine residues. They are generally associated with repression of expression.
Citrullination is the deimination of arginine residues, which leads to conversion of arginine to citrulline. This neutralizes the positive charge in the residue, and this reaction is catalyzed by a family of enzymes called protein arginine deiminases (PADs). This modification has been associated with both, repressive and activating functions.
O-GlcNAc glycosylationinvolves addition of N-acetylglucosamine (O-GlcNAc) sugar residues on the serine and threonine amino acid side chains, by O-GlcNAc transferase (OGT) using UDP-GlcNAc, as sugar donor. Removal of the sugar is catalyzed by the N-acetylglucosaminidase (O-GlcNAcase). These modifications can be either repressive or activating marks.
Post-translational modifications at the histone cores
Although most gene-regulating histone modifications occur at the N-terminal histone tails, there are some modifications also to the core domains of the histone proteins. They also can influence chromatin structure and gene expression. Some important core modifications include histone H3K56 acetylation, histone H3K79 methylation, histone H3T118 phosphorylation, histone H3K122 acetylation, histone H4K59 methylation, histone H4K91 acetylation, and histone H2AK99 methylation among others.