N6-Methyladenosine, or m6A, is formed when a methyl group is chemically added at the nitrogen-6 position of adenosine (A) residues. Often referred to as “the fifth RNA base”, m6A is the most common and abundant eukaryotic RNA modification, accounting for over 80% of all RNA methylation. It can be found mainly in mRNA, but is also observed in non-coding species (tRNA, rRNA, miRNA).
Formation and function of RNA methylation
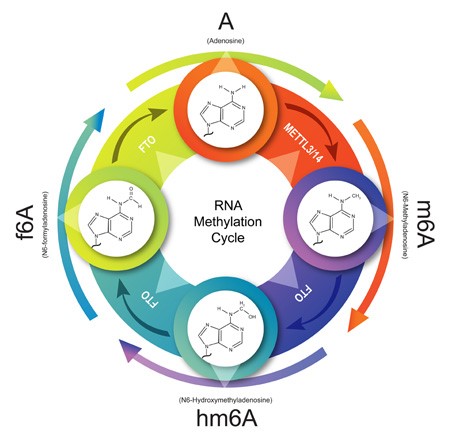
The formation of m6A RNA appears to be a co-transcriptional event taking place early on in the RNA lifecycle and is mediated by a multi-protein methyltransferase complex composed, in part, of METTL enzymes. Chemical addition of a methyl group to form m6A occurs via the METTL3/14 heterodimer. Through FTO-mediated oxidative demethylation, m6A is converted in a step-wise manner to hm6A and subsequently f6A before finally reverting back to A (Figure 1).
Figure 1. The m6A RNA cycle.
Through interactions with various binding proteins called “readers”, m6A affects virtually every facet of ribonucleic acid biology: structure, splicing, localization, translation, stability, and turnover [1]. Aside from this central role in RNA metabolism, m6A is a factor in other physiological processes such as cell differentiation, immunity, inflammation, and the circadian clock [2].
m6A RNA, disease, and therapeutic targets
The recent discoveries of these methylase “writers” (METTL3/14, WTAP, RBM15/15B, KIAA1429) and their associated demethylase “erasers” (FTO, ALKBH5, TET-like enzymes) in mammals uncovered the reversibility of the m6A modification, revealing potential therapeutic targets for m6A dysregulation-related diseases. Abnormal m6A methylation is implicated in diverse pathologies, although its role in disease has been most extensively investigated in the context of cancer.
Writers, erasers, and readers of m6A RNA methylation alike are linked to a wide array of oncological indications. METTL3 and METTL14 are reported in the pathogenesis of bladder and uterine cancer [3], endometrial cancer [4], renal cell carcinoma [5], acute myeloid leukemia (AML) [6-8], glioblastoma [9], and hepatocellular carcinoma [10,11]. FTO’s involvement in AML [12-15] and melanoma [16] has been documented, as well as ALKBH5’s connection to breast cancer [17] and glioblastoma [18]. An association between the m6A reader YTHDF2 and AML has also been established [19,20]. In addition to cancer, m6A writers and erasers have been studied in other diseases, including heart failure [21,22] and type 2 diabetes [23,24].
Effects of m6A on viral infection
Epigenetic modifications are known to influence the life cycles of RNA viruses like human coronavirus (CoV). Modified adenosines like m6A are reported to affect the viability of specific RNA viruses by modulating viral cap structures, viral replication, innate sensing pathways, and the innate immune response [25]. Interestingly, m6A exhibits both pro- and anti-viral activities, depending on the virus species and host cell type [26]. This makes the viral epitranscriptome an attractive target for remedial intervention.
Members of the coronaviruses as well as non-CoV species encode their own methyltransferases for self-methylating A residues and promoting immune evasion. Thus, examining the effects of m6A methylase/demethylase activity on the epigenome and epitranscriptome of coronaviruses and other RNA viruses, along with their targets, may provide valuable insights into the impact of viral and host-derived m6A RNA modifications on infection so as to guide remedial courses of action.
The RNA genome of SARS-CoV-2, the coronavirus responsible for the present COVID-19 pandemic, for example, contains more than 50 potential m6A sites based on the presence of specific sequence motifs for m6A modification by METTL3/14, including GGACU(T), GGACA, and GGACC. Consequently, greater than 0.64% of all adenosines, or 0.18% of all bases, in SARS-CoV-2 RNA could be m6A. Gain or loss of m6A can result in significant functional changes to RNA viruses, altering host cell fusion/entry, replication, transmission, pathogen intensity, and immune evasion. Results from recent METTL3 inhibitor and depletion studies demonstrating epigenomic suppression of SARS-CoV-2 are indeed encouraging, and suggest m6A methylase as a viable druggable target for remedial intervention. Knockdown of host cell METTL3 was shown to decrease SARS-CoV-2 m6A RNA methylation, viral load, percentage of infected cells, infectious virus production, and gene expression of proviral host factors such as ACE2 and TMPRSS2 [27,28]. The reduction of viral genome m6A levels enhanced viral recognition by the innate immune system (RIG-I binding to SARS-CoV-2 RNA), subsequently activating downstream innate immune signaling pathways and inflammatory gene expression [27].
What’s next?
Reliable assays for the detection of the m6A RNA modification and its modifying enzymes would undoubtedly benefit cancer and virus infection research and the development of novel therapeutics. Whether it’s quantifying m6A globally, assessing activity levels of m6A methyltransferases and demethylases, or high throughput screening of drugs directed at these epigenetic targets, EpigenTek’s portfolio of ELISA-based products offers a simple, convenient, and affordable means to pursue your RNA methylation research needs. Our proprietary EpiQuik and Epigenase technologies provide for rapid and direct measurements of N6-methyladenosine in RNA and activity/inhibition of m6A methylating and demethylating enzymes.
Hastings MH. m(6)A mRNA methylation: a new circadian pacesetter. Cell. 2013;155(4):740-741. doi:10.1016/j.cell.2013.10.028
Zhao S, Liu J, Nanga P, et al. Detailed modeling of positive selection improves detection of cancer driver genes. Nat Commun. 2019;10(1):3399. Published 2019 Jul 30. doi:10.1038/s41467-019-11284-9
Liu J, Eckert MA, Harada BT, et al. m6A mRNA methylation regulates AKT activity to promote the proliferation and tumorigenicity of endometrial cancer. Nat Cell Biol. 2018;20(9):1074-1083. doi:10.1038/s41556-018-0174-4
Li X, Tang J, Huang W, et al. The M6A methyltransferase METTL3: acting as a tumor suppressor in renal cell carcinoma. Oncotarget. 2017;8(56):96103-96116. Published 2017 Oct 10. doi:10.18632/oncotarget.21726
Barbieri I, Tzelepis K, Pandolfini L, et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature. 2017;552(7683):126-131. doi:10.1038/nature24678
Vu LP, Pickering BF, Cheng Y, et al. The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat Med. 2017;23(11):1369-1376. doi:10.1038/nm.4416
Weng H, Huang H, Wu H, et al. METTL14 Inhibits Hematopoietic Stem/Progenitor Differentiation and Promotes Leukemogenesis via mRNA m6A Modification. Cell Stem Cell. 2018;22(2):191-205.e9. doi:10.1016/j.stem.2017.11.016
Cui Q, Shi H, Ye P, et al. m6A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017;18(11):2622-2634. doi:10.1016/j.celrep.2017.02.059
Ma JZ, Yang F, Zhou CC, et al. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N6 -methyladenosine-dependent primary MicroRNA processing. Hepatology. 2017;65(2):529-543. doi:10.1002/hep.28885
Chen M, Wei L, Law CT, et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology. 2018;67(6):2254-2270. doi:10.1002/hep.29683
Li Z, Weng H, Su R, et al. FTO Plays an Oncogenic Role in Acute Myeloid Leukemia as a N6-Methyladenosine RNA Demethylase. Cancer Cell. 2017;31(1):127-141. doi:10.1016/j.ccell.2016.11.017
Su R, Dong L, Li C, et al. R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling. Cell. 2018;172(1-2):90-105.e23. doi:10.1016/j.cell.2017.11.031
Yan F, Al-Kali A, Zhang Z, et al. A dynamic N6-methyladenosine methylome regulates intrinsic and acquired resistance to tyrosine kinase inhibitors. Cell Res. 2018;28(11):1062-1076. doi:10.1038/s41422-018-0097-4
Huang Y, Su R, Sheng Y, et al. Small-Molecule Targeting of Oncogenic FTO Demethylase in Acute Myeloid Leukemia. Cancer Cell. 2019;35(4):677-691.e10. doi:10.1016/j.ccell.2019.03.006
Yang S, Wei J, Cui YH, et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat Commun. 2019;10(1):2782. Published 2019 Jun 25. doi:10.1038/s41467-019-10669-0
Zhang C, Samanta D, Lu H, et al. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m⁶A-demethylation of NANOG mRNA. Proc Natl Acad Sci U S A. 2016;113(14):E2047-E2056. doi:10.1073/pnas.1602883113
Zhang S, Zhao BS, Zhou A, et al. m6A Demethylase ALKBH5 Maintains Tumorigenicity of Glioblastoma Stem-like Cells by Sustaining FOXM1 Expression and Cell Proliferation Program. Cancer Cell. 2017;31(4):591-606.e6. doi:10.1016/j.ccell.2017.02.013
Wang H, Zuo H, Liu J, et al. Loss of YTHDF2-mediated m6A-dependent mRNA clearance facilitates hematopoietic stem cell regeneration. Cell Res. 2018;28(10):1035-1038. doi:10.1038/s41422-018-0082-y
Paris J, Morgan M, Campos J, et al. Targeting the RNA m6A Reader YTHDF2 Selectively Compromises Cancer Stem Cells in Acute Myeloid Leukemia. Cell Stem Cell. 2019;25(1):137-148.e6. doi:10.1016/j.stem.2019.03.021
Berulava T, Buchholz E, Elerdashvili V, et al. Changes in m6A RNA methylation contribute to heart failure progression by modulating translation. Eur J Heart Fail. 2020;22(1):54-66. doi:10.1002/ejhf.1672
Mathiyalagan P, Adamiak M, Mayourian J, et al. FTO-Dependent N6-Methyladenosine Regulates Cardiac Function During Remodeling and Repair. Circulation. 2019;139(4):518-532. doi:10.1161/CIRCULATIONAHA.118.033794
De Jesus DF, Zhang Z, Kahraman S, et al. m6A mRNA Methylation Regulates Human β-Cell Biology in Physiological States and in Type 2 Diabetes. Nat Metab. 2019;1(8):765-774. doi:10.1038/s42255-019-0089-9
Yang Y, Shen F, Huang W, et al. Glucose Is Involved in the Dynamic Regulation of m6A in Patients With Type 2 Diabetes. J Clin Endocrinol Metab. 2019;104(3):665-673. doi:10.1210/jc.2018-00619
Gonzales-van Horn SR, Sarnow P. Making the Mark: The Role of Adenosine Modifications in the Life Cycle of RNA Viruses. Cell Host Microbe. 2017;21(6):661-669. doi:10.1016/j.chom.2017.05.008
Dang W, Xie Y, Cao P, et al. N6-Methyladenosine and Viral Infection. Front Microbiol. 2019;10:417. Published 2019 Mar 5. doi:10.3389/fmicb.2019.00417
Li N, Hui H, Bray B, et al. METTL3 regulates viral m6A RNA modification and host cell innate immune responses during SARS-CoV-2 infection. Cell Rep. 2021;35(6):109091. doi:10.1016/j.celrep.2021.109091
Burgess HM, Depledge DP, Thompson L, et al. Targeting the m6A RNA modification pathway blocks SARS-CoV-2 and HCoV-OC43 replication. Genes Dev. 2021;35(13-14):1005-1019. doi:10.1101/gad.348320.121





 Cart (0)
Cart (0)