Choose the catalog number and quantity to pre-fill on the quote request page.
MethylFlash Hydroxymethylated DNA 5-hmC Quantification Kit (Colorimetric)
FTL Polyclonal Antibody, FITC Conjugated
EpiQuik Hydroxymethylated DNA Immunoprecipitation (hMeDIP) Kit
5-Hydroxymethylcytosine (5-hmC) Monoclonal Antibody [HMC/4D9]
Epigenase 5mC-Hydroxylase TET Activity/Inhibition Assay Kit (Colorimetric)
FTL Polyclonal Antibody
We recommend using a newer version of this product: MethylFlash Global DNA Hydroxymethylation (5-hmC) ELISA Easy Kit (Colorimetric)
The MethylFlash™ Hydroxymethylated DNA Quantification Kit (Colorimetric) is a complete set of optimized buffers and reagents to colorimetrically quantify global DNA hydroxymethylation by specifically measuring levels of 5-hydroxymethylcytosine (5-hmC) without cross-reactivity to methylcytosine and unmethylated cytosine, in an ELISA-like microplate-based format.
This kit is also specifically optimized for paired use with the MethylFlash Methylated DNA Quantification Kit (Colorimetric) for simultaneously quantifying both methylated DNA and hydroxymethylated DNA.
About 5-Hydroxymethylcytosine5-hmC is a modified form of cytosine, recently discovered in animal tissues. The function of 5-hmC in epigenetics may be different from its forerunner 5-methylcytosine (5-mC) and currently remains a mystery. It is believed, though, that 5-hmC plays an important role in switching genes on and off. The presence of 5-hmC makes it necessary to not only re-evaluate existing DNA methylation data, but also necessary to determine the relative distribution and changes of 5-hmC in human tissues of healthy and diseased statuses. Prior to Epigentek’s MethylFlash™ technology, there were no methods that could be used for practically and routinely identifying 5-hmC and discriminating this base from 5-mC.
Distinguishing Between 5-hmC and 5-mCCurrently used methylated DNA analysis methods including restriction enzyme digestion and bisulfite or MeDIP-mediated MS-PCR and sequencing are not suitable for 5-hmC detection as 5-hmC and 5-mC are virtually indistinguishable between each other with these methods. To address this problem, Epigentek offers the MethylFlash™ Hydroxymethylated DNA Quantification Kit which uses a unique procedure to quantify global DNA hydroxymethylation. This product provides a cost-effective way to measure levels of 5-hydroxymethylcytosine and to distinguish between 5hmC, 5mC, and C. This allows for researchers to re-evaluate their DNA methylation data for DNA hydroxymethylation and to efficiently look for DNA hydroxymethylation in new DNA samples. The kit has the following advantages and features:
Comparison of Available ELISA-based Global DNA Methylation Assay Methods
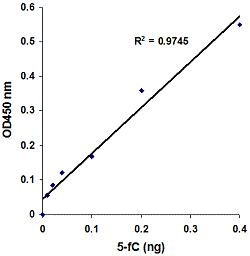
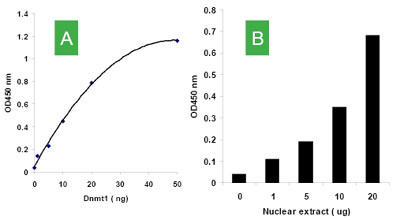
Principle & ProcedureThe MethylFlash™ Hydroxymethylated DNA Quantification Kit (Colorimetric) contains all reagents necessary for the colorimetric quantification of global DNA hydroxymethylation. In this assay, DNA is bound to strip wells that are specifically treated to have a high DNA affinity. The hydroxymethylated fraction of DNA is detected using capture and detection antibodies and then quantified through an ELISA-like reaction by reading the absorbance in a microplate spectrophotometer at 450 nm. The amount of hydroxymethylated DNA is proportional to the OD intensity measured, which can be calculated with the kit's included formulas for relative hydroxymethylation status of two different DNA samples or absolute quantification of 5-hydroxymethylcytosine (5-hmC) using a standard curve.
Easy, Fast, and FlexibleThe entire colorimetric assay has easy-to-follow steps for convenience and speed, which can be completed in 3 hours and 45 minutes. The strip-well microplate format allows for a flexible assay in manual or high throughput analysis. Universal positive and negative controls are included with the kit for quantifying hydroxymethylated DNA from any species such as mammals, plants, fungi, bacteria, and viruses.
Safe and ConvenientAll the needed reagents, including negative controls and positive controls, for quantification of global DNA hydroxymethylation are conveniently packaged in the kit. The direct colorimetric quantification of DNA samples replaces obsolete or inferior methods and eliminates the need for DNA digestion/denaturation, radioactivity, extraction, or chromatography.
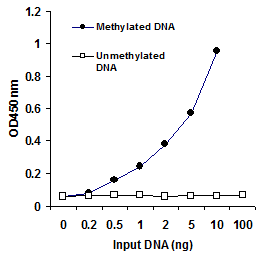
Highly Sensitive and SpecificThe novel procedure and proprietary kit compositions allow for accurate quantification of hydroxymethylated DNA to be achieved with high sensitivity and specificity. The detection limit of the input DNA can be as low as 40 pg of hydroxymethylated DNA with a dynamic range from 40 pg to 20 ng (see Fig. 2). Analytic sensitivity is about ten-fold higher than that obtained by HPLC (0.008% vs 0.08%). There is no cross-reactivity to methylcytosine and unmethylated cytosine so that only hydroxymethylated DNA (5-hmC) is detected (see Fig. 3).
Responsive, Reliable, and PracticalBased on its working principle and the microplate format, the kit can be practically and routinely used for any species from a variety of forms including from cultured cells, fresh and frozen tissues, paraffin-embedded tissues, plasma/serum samples, and body fluid samples. To demonstrate the capabilities of the kit, it has been successfully used for quantifying the content of 5-hmC in DNA from mouse brains and HeLa cells. The percentage of 5-hmC measured by the MethylFlash™ kit is parallel and comparable to that detected by HPLC and LC-MS methods (see Table 1). Through the use of the kit, a high abundance of 5-hmC in normal human brains and colon tissues is discovered for the first time. Furthermore, it shows that 5-hmC accounts for 32% and 16% of total methylated cytosine (5-mC plus 5-hmC) in normal human brains and colon tissues, respectively (see Table 2). In contrast, 5-hmC is low or absent in colon cancer cell lines (HCT116) and cervical cancer cell lines (HeLa) (see Fig. 4A and 4B).
Table 1. Comparison of the MethylFlash™ Hydroxymethylated DNA Quantification Kit and chromatography-based methods for the detection of 5-hydroxymethylcytosine, shown in percentage of 5-hmC present in two DNA sample types.
1 Munzel M et al: Angewandte Chemie, 122: ePub, 2010
Table 2. Percentage of 5-hmC and 5-mC in different tissues/cells, measured with the MethylFlash™ Hydroxymethylated DNA Quantification Kit and the MethylFlash™ Methylated DNA Quantification Kit, respectively.
Fig. 4. Demonstration of a high abundance of 5-hydroxymethylcytosine in human brain and colon tissues. 200 ng of DNA isolated from different tissues or cells were added into the assay wells and the hydroxymethylated fraction of DNA was measured using the MethylFlash™ Hydroxymethylated DNA Quantification Kit. A: OD intensity measured in variety of DNA samples. B: Percentage of hydroxymethylated DNA in various tissues and cells.





 Cart (0)
Cart (0)


![5-Hydroxymethylcytosine (5-hmC) Monoclonal Antibody [HMC/4D9]](images/a1018data.gif)