The Discovery and Prevalence of N6-methyladenosine (m6A)
First discovered in the 1970s, N6-methyladenosine, or m6A, remains the most extensively studied RNA modification. As the most prolific methylation modification in eukaryotic mRNA, m6A has been observed in a wide array of organisms: yeast, plants, insects, mammals, and even viruses. In mammals, this epigenetic modification is present in many different tissues, with the highest expression in the brain, kidney, and liver. It is involved in diverse physiologic processes, including stem cell differentiation, cell division, gametogenesis, and biological rhythms, while aberrancies in m6A methylation have been implicated in assorted complications such as tumors, obesity, and infertility.

Advancements in Understanding m6A Methylation
It was not until a little over a decade ago that scientists gained a better understanding of the modification’s exact location, how it changes over time, and how it is controlled. This progress was forged ahead by the discovery of the first RNA demethylase, fat mass and obesity-associated (FTO) protein [Jia 2011], along with technological advancements like methylated RNA immunoprecipitation and high-throughput sequencing that allowed researchers to accurately map the distribution of m6A across the entire transcriptome.
Addition, deletion, and functionality of the m6A RNA modification
m6A is formed when a methyl group is chemically added at the nitrogen-6 position of adenosine residues. It is a reversible modification that occurs primarily on the A of a highly conserved mRNA sequence known as the RRACH motif, where R can be either G or A (with a preference for G), and H can be U, A, or C (with a preference for U) [Harper 1990]. m6A RNA methylation is regulated by three main types of biomolecules: methyltransferases, or “writers”; demethylases, or “erasers”; and binding proteins, or “readers”, which interact with the m6A-modified RNA to elicit a range of biological functions (Figure 1).

m6A Writers
The methylase that catalyzes m6A RNA formation is a multicomponent complex comprised of the core subunits methyltransferase like 3 (METTL3), METTL14, and Wilms tumor 1-associating protein (WTAP), as well as supplementary components that include vir like m6A methyltransferase associated (VIRMA), RNA binding motif protein 15 (RBM15), and RBM15B. METTL3 can bind to the methyl group donor S-adenosylmethionine (SAM) and catalyze the formation of m6A [Bokar 1997]. It forms a stable complex with METTL14, and the resulting heterodimer can subsequently engage WTAP, which localizes the methyltransferase complex in the nucleus [Ping 2014]. VIRMA, RBM15, and RBM15B have all been reported to interact with WTAP and recruit the complex to target RNAs [Patil 2016; Yue 2018].
m6A Erasers
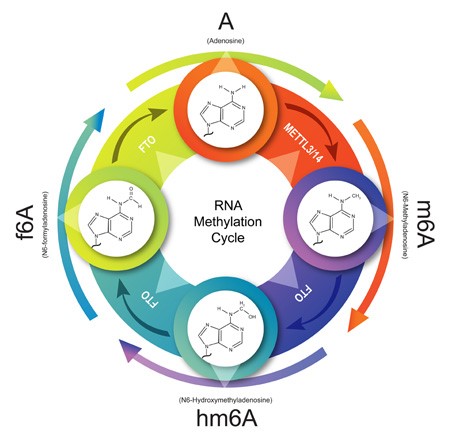
The m6A demethylases FTO and alkB homolog 5 (ALKBH5) [Zheng 2013] can reverse the m6A methylation process by removing the methyl group from the RNA molecule. Through FTO-mediated oxidative demethylation, m6A is converted in a step-wise manner to N6-hydroximethyladenosine (hm6A) and subsequently N6-formyldenosine (f6A) before finally reverting back to A (Figure 2) [Fu 2013]. Whether such intermediates are generated during m6A demethylation by ALKBH5 requires further investigation [Toh 2020].
m6A Readers
The biofunction of m6A primarily involves post-transcriptional regulation of RNA though interactions with m6A-binding proteins. Of particular note are the YT521-B homology (YTH) N6-methyladenosine RNA binding proteins (YTHDF1-3, YTHDC1-2) and the heterogeneous nuclear ribonucleoproteins (HNRNPA2B1, HNRNPC, HNRNPG). Researchers have discovered that, via these so-called “readers”, m6A affects virtually every facet of ribonucleic acid biology: structure, splicing, localization, translation, stability, and turnover [Zaccara 2019].
YTHDF1 promotes translation of m6A-mRNA by facilitating RNA binding to ribosomes [Wang 2015]. YTHDF2 mediates the degradation of m6A-modified RNAs, including mRNA and some long non-coding RNA [Wang 2014]. YTHDF3 enhances interaction of YTHDF1 and YTHDF2 to their respective RNA substrates, thereby fostering protein synthesis or RNA decay [Shi 2017]. YTHDC1 modulates alternative splicing and nuclear export of m6A-containing mRNA [Xiao 2016; Roundtree 2017], while YTHDC2 enriches the translation efficiency of modified transcripts. HNRNPA2B1, HNRNPC, and HNRNPG have all been identified as regulators of alternative splicing in an m6A-dependent manner [Alarcón 2015; Liu 2015; Liu 2017]. Aside from this central role in RNA metabolism, m6A is a factor in other physiological processes such as cell differentiation, immunity, inflammation, and the circadian clock [Hasting 2013]. Abnormal m6A methylation has been implicated in diverse pathologies: diabetes, obesity, neurodegeneration, and cancer, to name a few. The recent discoveries of m6A methylase “writers” and their associated demethylase “erasers” in mammals uncovered the reversibility of the m6A modification, exposing potential therapeutic targets for m6A dysregulation-related diseases. Continued research in elucidating the dynamics of the m6A RNA methylation machinery, its various components, and the interplay among those components will undoubtedly support the design and development of novel therapies.
Related Products
- P-9005 EpiQuik™ m6A RNA Methylation Quantification Kit (Colorimetric)
- P-9018 EpiQuik™ CUT&RUN m6A RNA Enrichment (MeRIP) Kit
- P-9013 Epigenase™ m6A Demethylase Activity/Inhibition Assay Kit (Colorimetric)
- P-9019 Epigenase™ m6A Methylase Activity/Inhibition Assay Kit (Colorimetric)
References
- Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell. 2015;162(6):1299-1308. doi:10.1016/j.cell.2015.08.011
- Bokar JA, Shambaugh ME, Polayes D, Matera AG, Rottman FM. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. RNA. 1997;3(11):1233-1247.
- Fu Y, Jia G, Pang X, et al. FTO-mediated formation of N6-hydroxymethyladenosine and N6-formyladenosine in mammalian RNA. Nat Commun. 2013;4:1798. doi:10.1038/ncomms2822
- Harper JE, Miceli SM, Roberts RJ, Manley JL. Sequence specificity of the human mRNA N6-adenosine methylase in vitro. Nucleic Acids Res. 1990;18(19):5735-5741. doi:10.1093/nar/18.19.5735
- Hastings MH. m(6)A mRNA methylation: a new circadian pacesetter. Cell. 2013;155(4):740-741. doi:10.1016/j.cell.2013.10.028
- Jia G, Fu Y, Zhao X, et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO [published correction appears in Nat Chem Biol. 2012 Dec;8(12):1008]. Nat Chem Biol. 2011;7(12):885-887. Published 2011 Oct 16. doi:10.1038/nchembio.687
- Liu N, Dai Q, Zheng G, He C, Parisien M, Pan T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature. 2015;518(7540):560-564. doi:10.1038/nature14234
- Liu N, Zhou KI, Parisien M, Dai Q, Diatchenko L, Pan T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017;45(10):6051-6063. doi:10.1093/nar/gkx141
- Patil DP, Chen CK, Pickering BF, et al. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature. 2016;537(7620):369-373. doi:10.1038/nature19342
- Ping XL, Sun BF, Wang L, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014;24(2):177-189. doi:10.1038/cr.2014.3
- Roundtree IA, Luo GZ, Zhang Z, et al. YTHDC1 mediates nuclear export of N6-methyladenosine methylated mRNAs. Elife. 2017;6:e31311. Published 2017 Oct 6. doi:10.7554/eLife.31311
- Shi H, Wang X, Lu Z, et al. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017;27(3):315-328. doi:10.1038/cr.2017.15
- Toh JDW, Crossley SWM, Bruemmer KJ, et al. Distinct RNA N-demethylation pathways catalyzed by nonheme iron ALKBH5 and FTO enzymes enable regulation of formaldehyde release rates. Proc Natl Acad Sci U S A. 2020;117(41):25284-25292. doi:10.1073/pnas.2007349117
- Wang X, Lu Z, Gomez A, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505(7481):117-120. doi:10.1038/nature12730
- Wang X, Zhao BS, Roundtree IA, et al. N(6)-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell. 2015;161(6):1388-1399. doi:10.1016/j.cell.2015.05.014
- Xiao W, Adhikari S, Dahal U, et al. Nuclear m(6)A Reader YTHDC1 Regulates mRNA Splicing [published correction appears in Mol Cell. 2016 Mar 17;61(6):925]. Mol Cell. 2016;61(4):507-519. doi:10.1016/j.molcel.2016.01.012
- Yue Y, Liu J, Cui X, et al. VIRMA mediates preferential m6A mRNA methylation in 3'UTR and near stop codon and associates with alternative polyadenylation. Cell Discov. 2018;4:10. Published 2018 Feb 27. doi:10.1038/s41421-018-0019-0
- Zaccara S, Ries RJ, Jaffrey SR. Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol. 2019;20(10):608-624. doi:10.1038/s41580-019-0168-5
- Zheng G, Dahl JA, Niu Y, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. 2013;49(1):18-29. doi:10.1016/j.molcel.2012.10.015