Choose the catalog number and quantity to pre-fill on the quote request page.
EpiQuik Histone Methyltransferase Activity/Inhibition Assay Kit (H3K27)
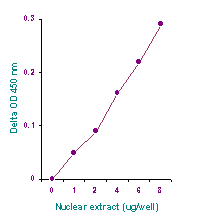
EpiQuik Nuclear Extraction Kit
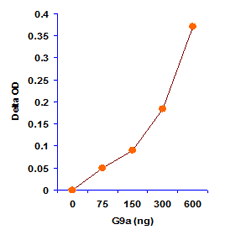
EpiQuik Histone Methyltransferase Activity/Inhibition Assay Kit (H3K9)
EpiQuik DNA Methyltransferase (DNMT) Activity/Inhibition Assay Kit
Histone H4K79me2 (H4K79 Dimethyl) Polyclonal Antibody
MUC2 Polyclonal Antibody, Biotin Conjugated
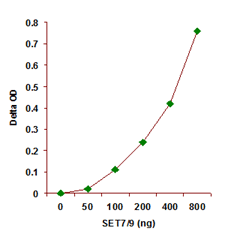
The EpiQuik™ Histone Methyltransferase Activity/Inhibition Assay Kit (H3-K27) is a convenient set of tools that allows the experimenter to measure the activity or inhibition of individual histone methyltransferase that specifically target histone H3 at lysine 27 (H3-K27). The kit is ready-to-use and provides all the essential components needed to carry out a successful HMT activity/inhibition experiment without the need for radioactivity or any special equipment. The kit has the following advantages:
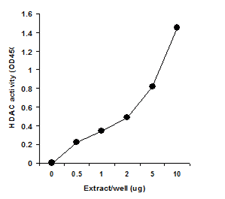
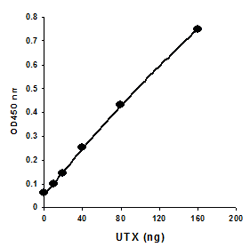
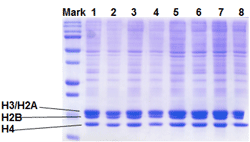
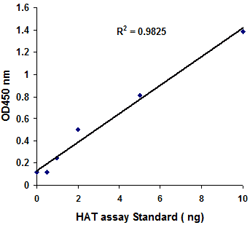
Principle & ProcedureThe EpiQuik™ Histone Methyltransferase Activity/Inhibition Assay Kit (H3-K27) is designed for measuring HMTs that specifically target histone H3 at lysine 27. In an assay with this kit, the histone substrate is stably captured on the strip wells through biotin-streptavidin binding. HMT enzymes transfer a methyl group to histone H3 substrate from Adomet to methylate the substrate at lysine 4. The methylated histone H3-K27 can be recognized with a high-affinity antibody. The ratio or amount of methylated H3- K27, which is directly proportional to enzyme activity, can be quantified through a HRP conjugated secondary antibody-color development system. The HMT activity is then calculated based on the amount of methylated H3-K27 converted by the HMTs.





 Cart (0)
Cart (0)