N6-methyladenosine, or m6A, is the most common and abundant eukaryotic RNA modification, accounting for over 80% of all RNA methylation. It can be found mainly in mRNA, but is also observed in non-coding species like tRNA, rRNA, and miRNA. Through interactions with various binding proteins called “readers”, m6A affects virtually every facet of ribonucleic acid biology: structure, splicing, localization, translation, stability, and turnover [1]. Aside from this central role in RNA metabolism, m6A is a factor in other physiological processes such as cell differentiation, immunity, inflammation, and the circadian clock [2]. Abnormal m6A methylation has been implicated in diverse pathologies: diabetes, obesity, neurodegeneration, and cancer, to name a few. The formation of m6A RNA appears to be a co-transcriptional event occurring early on in the RNA lifecycle and is mediated by a multi-protein methyltransferase complex.
Mapping the m6A epitranscriptome
Several methods are available for epitranscriptome-wide m6A mapping:
MeRIP-seq.
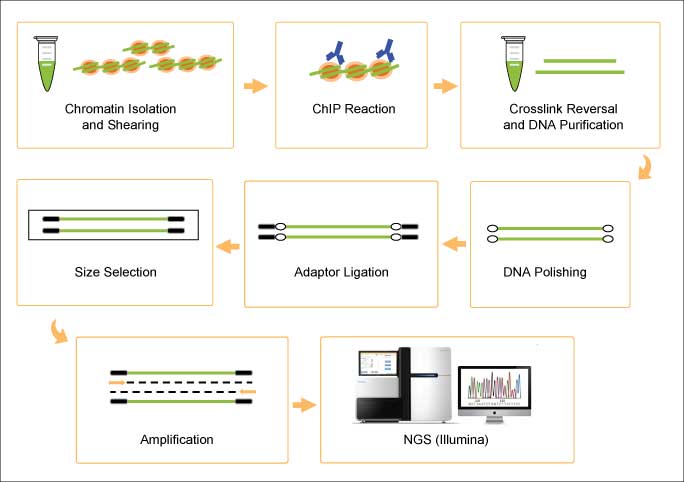
The development of methylated RNA immunoprecipitation sequencing (MeRIP-seq) was a landmark in the field of epitranscriptomics as it was the first method to detect m6A on a transcriptome-wide level [3]. MeRIP-seq couples m6A RNA immunoprecipitation with NGS, providing for the high-throughput localization of modified sites from enriched m6A-containing RNA fragments that have been precipitated by a specific antibody, reverse-transcribed, and sequenced. The fragment sizes generated during the random fragmentation step preceding immunoprecipitation limit the precise mapping of the m6A site to within a ~200 nt stretch. The choice of antibody allows MeRIP-seq to be adapted toward the study of other modified RNA types (e.g., 5hmC RNA [4]).
Schematic of the MeRIP-seq protocol [3].
miCLIP.
m6A individual-nucleotide resolution crosslinking and immunoprecipitation (miCLIP) was developed to address the disadvantages associated with MeRIP-seq approaches regarding the mapping of m6A RNA sites at individual-nucleotide resolution [5]. The key feature of this method is the UV-crosslinking of immunoprecipitated RNA fragments to the capture antibody. Antibody remnants at the crosslinking site on the RNA after Proteinase K treatment induce signature mutations (truncations and C→T transitions) during cDNA synthesis that can be identified by sequencing and used to more precisely map the specific m6A location. These antibody-induced mutational signatures have also been successfully applied to the mapping of m6Am RNA modifications.
PA-m6A-seq.
Photo-crosslinking-assisted m6A sequencing (PA-m6A-seq) is an alternative UV-based strategy that was fashioned for high-resolution (~23 nts) transcriptome-wide m6A mapping [6]. This method employs a photoreactive ribonucleoside crosslinker to induce a signature mutation for localizing m6A, akin to miCLIP. The uridine analogue 4-thiouridine (4SU) is incorporated into sample RNA. Full-length, unfragmented, 4SU-labeled RNA molecules are then immunoprecipitated with an anti-m6A antibody, and UV irradiation is applied to establish covalent crosslinks between m6A-bound antibody and neighboring 4SU. Crosslinked RNA is digested with RNase T1 to yield ~30 nt-long fragments that are further processed for library preparation and sequencing, whereby crosslink-generated T→C transitions adjacent to the m6A sites can be identified.
eTAM-seq.
Evolved TadA-assisted N6-methyladenosine sequencing (eTAM-seq) is the latest addition to the researcher’s m6A profiling toolbox [7]. eTAM-seq detects and quantifies m6A via global adenosine deamination, a process that converts all unmethylated A into I, which is subsequently read as G, while m6A remains unaffected. The A-to-I conversion is facilitated by a distinctive variant (TadA8.20) of the E. coli TadA enzyme that maintains RNA integrity and thus minimizes sample loss to a greater extent when compared with harsher chemical deamination. Given the novelty of this enzyme-assisted sequencing technology and lack of in-depth characterization, several concerns still require attention, including time-consuming protocol; sensitivity (eTAM-seq is less sensitive to lowly methylated A sites); enzyme accessibility (eTAM-seq may not be applicable for highly structured RNA, and has high sequence bias); enzyme specificity (the TadA8.20 deaminase may not convert non-m6A adenosine modifications); and high cost. eTAM-seq is currently not available commercially.
Cutting through the background noise
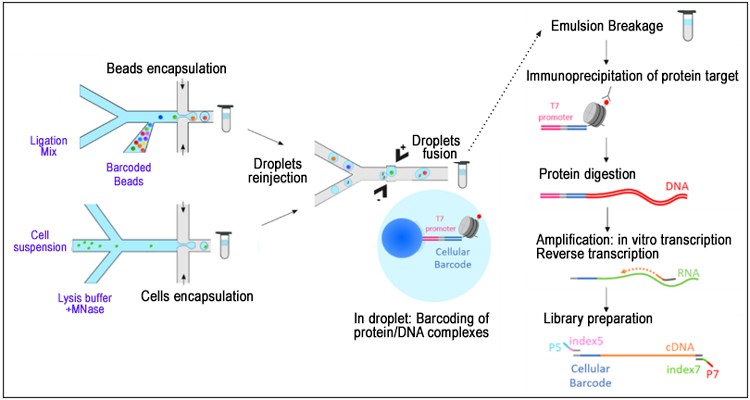
Cleavage under targets and release using nuclease (CUT&RUN) is a recent innovation in the study of protein/DNA interactions, designed to tackle the limitations of the popular ChIP-seq technique, namely, the need for a large amount of starting material, long protocol time, and limited resolution. This novel method, which is performed in situ on intact cells without fixation, entails cleavage of chromatin at specific sites by a unique fusion protein and the subsequent, direct capture of protein-DNA complexes. The fusion protein consists of: 1) a cleavage domain (e.g., micrococcal nuclease, or MNase); and 2) a protein A/G (pAG) domain that binds to an antibody against the protein of interest.
Schematic of the MeRIP-seq protocol [8].
During the procedure, the antibody specifically captures the target protein. The pAG-MNase fusion protein then binds to the antibody and cleaves the chromatin at the protein-DNA interaction site, resulting in the release of the protein-DNA complex from the chromatin. The CUT&RUN method eliminates interference from crosslinking and yields shorter-length DNA fragments, allowing for reduced background signals.
MeRIP with CUT&RUN for increased sensitivity
Established methods used for epitranscriptome-wide m6A mapping like MeRIP-seq, PA-m6A-seq, and miCLIP have either been widely used but are unable to achieve high resolution in m6A profiling; or improve the profiling resolution but suffer from poor reproducibility and a complicated process. In particular, they are time-consuming (>2 days) and costly.
To address these issues, EpigenTek has pioneered a new method: cleavage under target and recover using nuclease for m6A enrichment (CUT&RUN m6A MeRIP) [9-18]. This innovative approach, the first of its kind, combines the advantages of MeRIP-seq and miCLIP with CUT&RUN technology for higher enrichment, lower input, reduced background, and a faster, more streamlined procedure. CUT&RUN m6A MeRIP uses a state-of-the-art RNA cleavage enzyme mix to simultaneously fragment immunocaptured RNA and cleave/remove any RNA sequences in both ends of the target m6A-containing sequences without affecting RNA regions occupied by the antibody. Short RNA fragments are consequently generated only bound with anti-m6A antibody. True target m6A-enriched regions can therefore be reliably identified, and high-resolution mapping achieved.
The m6A peak distribution from samples processed with CUT&RUN m6A MeRIP correlates well with expected regions as shown in published data (Inset; see Ref 3).
Hastings MH. m(6)A mRNA methylation: a new circadian pacesetter. Cell. 2013;155(4):740-741. doi:10.1016/j.cell.2013.10.028
Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell. 2012;149(7):1635-1646. doi:10.1016/j.cell.2012.05.003
Delatte B, Wang F, Ngoc LV, et al. RNA biochemistry. Transcriptome-wide distribution and function of RNA hydroxymethylcytosine. Science. 2016;351(6270):282-285. doi:10.1126/science.aac5253
Linder B, Grozhik AV, Olarerin-George AO, Meydan C, Mason CE, Jaffrey SR. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods. 2015;12(8):767-772. doi:10.1038/nmeth.3453
Chen K, Lu Z, Wang X, et al. High-resolution N(6) -methyladenosine (m(6) A) map using photo-crosslinking-assisted m(6) A sequencing. Angew Chem Int Ed Engl. 2015;54(5):1587-1590. doi:10.1002/anie.201410647
Xiao YL, Liu S, Ge R, et al. Transcriptome-wide profiling and quantification of N6-methyladenosine by enzyme-assisted adenosine deamination [published online ahead of print, 2023 Jan 2]. Nat Biotechnol. 2023;10.1038/s41587-022-01587-6. doi:10.1038/s41587-022-01587-6
Kaya-Okur HS, Janssens DH, Henikoff JG, Ahmad K, Henikoff S. Efficient low-cost chromatin profiling with CUT&Tag. Nat Protoc. 2020;15(10):3264-3283. doi:10.1038/s41596-020-0373-x
Pan XY, Bi YH, Cheng M, et al. METTL3 facilitates hepatic fibrosis progression via m6A-YTHDF2 dependent silencing of GPR161. Preprint. bioRxiv. 2021; 2021.12.15.472749. Published 2021 Dec 15. doi:10.1101/2021.12.15.472749
Huang J, Sun W, Wang Z, et al. FTO suppresses glycolysis and growth of papillary thyroid cancer via decreasing stability of APOE mRNA in an N6-methyladenosine-dependent manner. J Exp Clin Cancer Res. 2022;41(1):42. Published 2022 Jan 28. doi:10.1186/s13046-022-02254-z
Zhou T, Chen G, Chen M, et al. Direct Full-Length RNA Sequencing Reveals an Important Role of Epigenetics During Sexual Reversal in Chinese Soft-Shelled Turtle. Front Cell Dev Biol. 2022;10:876045. Published 2022 Mar 25. doi:10.3389/fcell.2022.876045
Ji X, Sun W, Lv C, et al. ALKBH5-induced circRNA NRIP1 promotes glycolysis in thyroid cancer cells by targeting the miR-541-5p/PKM2 and miR-3064-5p/PKM2 axes. Preprint. Res Sq. 2022;rs.3.rs-1493404. Published 2022 Apr 4. doi:10.21203/rs.3.rs-1493404/v1
Chen YH, Jiang T, Fan BY, et al. Transcriptome-wide N6-methyladenosine methylome profiling of Bombyx mori reveals a potential mechanism of epigenetic regulation on diapause. Preprint. Res Sq. 2022;rs.3.rs-1561012. Published 2022 Apr 18. doi:10.21203/rs.3.rs-1561012/v1
Xu C, Huang H, Zhang M, et al. Methyltransferase-Like 3 Rescues the Amyloid-beta protein-Induced Reduction of Activity-Regulated Cytoskeleton Associated Protein Expression via YTHDF1-Dependent N6-Methyladenosine Modification. Front Aging Neurosci. 2022;14:890134. Published 2022 Apr 25. doi:10.3389/fnagi.2022.890134
Wang X, Lu X, Wang P, et al. SRSF9 promotes colorectal cancer progression via stabilizing DSN1 mRNA in an m6A-related manner. J Transl Med. 2022;20(1):198. Published 2022 May 4. doi:10.1186/s12967-022-03399-3





 Cart (0)
Cart (0)