How CUT&RUN Helps Improve Methylated RNA Immunoprecipitation
Often referred to as “the fifth RNA base”, N6-methyladenosine, or m6A, is the most common and abundant eukaryotic RNA modification, accounting for over 80% of all RNA methylation. It can be found mainly in mRNA, but is also observed in non-coding species like tRNA, rRNA, and miRNA. Through interactions with various binding proteins called “readers”, m6A affects virtually every facet of ribonucleic acid biology: structure, splicing, localization, translation, stability, and turnover [1]. Aside from this central role in RNA metabolism, m6A is a factor in other physiological processes such as cell differentiation, immunity, inflammation, and the circadian clock [2]. Abnormal m6A methylation has been implicated in diverse pathologies: diabetes, obesity, neurodegeneration, and cancer, to name a few. The formation of m6A RNA appears to be a co-transcriptional event occurring early on in the RNA lifecycle and is mediated by a multi-protein methyltransferase complex.
Currently, several methods are used for epitranscriptome-wide m6A mapping:
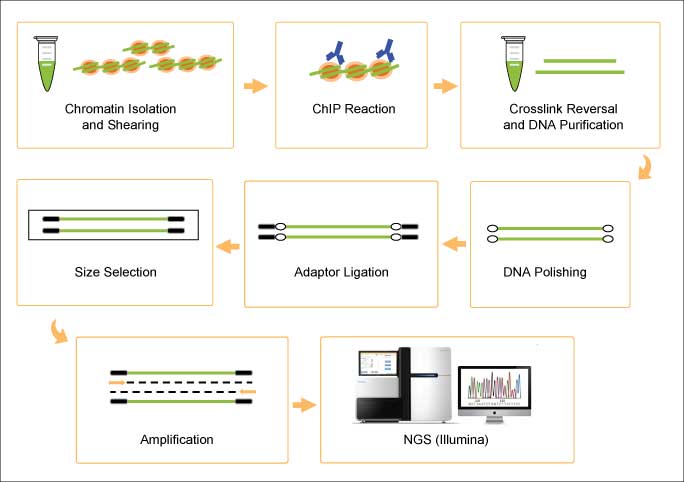
MeRIP-seq. The development of methylated RNA immunoprecipitation sequencing (MeRIP-seq) was a landmark in the field of epitranscriptomics as it was the first method to detect m6A on a transcriptome-wide level [3]. MeRIP-seq couples m6A RNA immunoprecipitation with NGS, allowing for the high-throughput localization of modified sites from enriched m6A-containing RNA fragments that have been precipitated by a specific antibody, reverse-transcribed, and sequenced. The fragment sizes generated during the random fragmentation step preceding immunoprecipitation limits the precise mapping of the m6A site to within a ~200 nt stretch. The choice of antibody allows MeRIP-seq to be adapted toward the study of other modified RNA types (e.g., 5hmC RNA, [4].
Schematic of the MeRIP-seq protocol as shown in Ref 3.
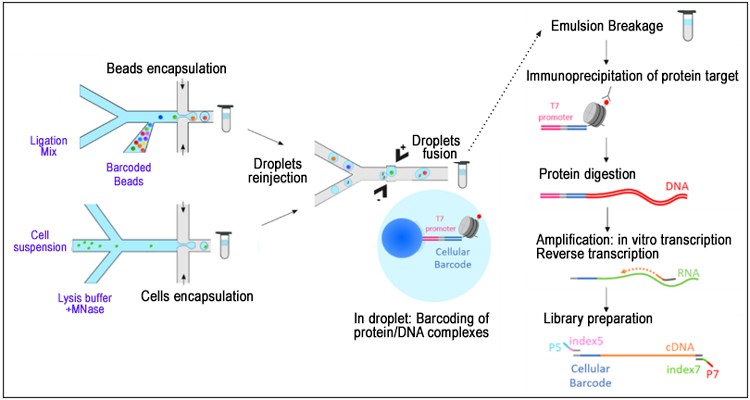
miCLIP. m6A individual-nucleotide resolution crosslinking and immunoprecipitation (miCLIP) was designed to address the disadvantages associated with MeRIP-seq approaches regarding the mapping of m6A RNA sites at individual-nucleotide resolution [5]. The key feature of this method is the UV-crosslinking of immunoprecipitated RNA fragments to the capture antibody. Antibody remnants at the crosslinking site on the RNA after Proteinase K treatment induce signature mutations (truncations and C→T transitions) during cDNA synthesis that can be identified by sequencing and used to more precisely map the specific m6A location. These antibody-induced mutational signatures have also been successfully applied to the mapping of m6Am RNA modifications.
Schematic of the miCLIP protocol as shown in Ref 5.
PA-m6A-seq. Photo-crosslinking-assisted m6A sequencing (PA-m6A-seq) is an alternative UV-based strategy that was fashioned for high-resolution (~23 nt) transcriptome-wide m6A mapping [6]. This method employs a photoreactive ribonucleoside crosslinker to induce a signature mutation for localizing m6A, akin to miCLIP. The uridine analogue 4-thiouridine (4SU) is incorporated into sample RNA. Full-length, unfragmented, 4SU-labeled RNA molecules are then immunoprecipitated with an anti-m6A antibody, and UV irradiation is applied to establish covalent crosslinks between m6A-bound antibody and neighboring 4SU. Crosslinked RNA is digested with RNase T1 to yield ~30 nt-long fragments that are further processed for library preparation and sequencing, whereby crosslink-generated T→C transitions adjacent to the m6A sites can be identified.
Schematic of the PA-m6A-seq protocol as shown in Ref 6.
Established methods used for epitranscriptome-wide m6A mapping like MeRIP-seq, PA-m6A-seq, and miCLIP have either been widely used but are unable to achieve high resolution in m6A profiling, or improve the profiling resolution but suffer from poor reproducibility and a complicated process. In particular, they are time-consuming (>2 days) and costly. To address these issues, EpigenTek has developed a new method: cleavage under target and recover using nuclease for m6A enrichment (CUT&RUN m6A MeRIP) [7-16]. This innovative approach combines the advantages of MeRIP-seq and miCLIP with EpigenTek’s proprietary EpiQuik technology for higher enrichment, lower input, reduced background, and a faster, more streamlined procedure. CUT&RUN m6A MeRIP uses a state-of-the-art RNA cleavage enzyme mix to simultaneously fragment immunocaptured RNA and cleave/remove any RNA sequences in both ends of the target m6A-containing sequences, without affecting RNA regions occupied by the antibody. Short RNA fragments are consequently generated only bound with anti-m6A antibody. True target m6A-enriched regions can therefore be reliably identified, and high-resolution mapping achieved.
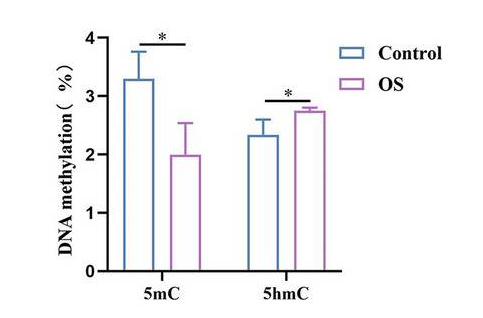
The m6A peak distribution from samples processed with CUT&RUN m6A MeRIP correlates well with expected regions as shown in published data (Inset; see Ref 3).
Hastings MH. m(6)A mRNA methylation: a new circadian pacesetter. Cell. 2013;155(4):740-741. doi:10.1016/j.cell.2013.10.028
Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell. 2012;149(7):1635-1646. doi:10.1016/j.cell.2012.05.003
Delatte B, Wang F, Ngoc LV, et al. RNA biochemistry. Transcriptome-wide distribution and function of RNA hydroxymethylcytosine. Science. 2016;351(6270):282-285. doi:10.1126/science.aac5253
Linder B, Grozhik AV, Olarerin-George AO, Meydan C, Mason CE, Jaffrey SR. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods. 2015;12(8):767-772. doi:10.1038/nmeth.3453
Chen K, Lu Z, Wang X, et al. High-resolution N(6) -methyladenosine (m(6) A) map using photo-crosslinking-assisted m(6) A sequencing. Angew Chem Int Ed Engl. 2015;54(5):1587-1590. doi:10.1002/anie.201410647
Pan XY, Bi YH, Cheng M, et al. METTL3 facilitates hepatic fibrosis progression via m6A-YTHDF2 dependent silencing of GPR161. Preprint. bioRxiv. 2021; 2021.12.15.472749. Published 2021 Dec 15. doi:10.1101/2021.12.15.472749
Huang J, Sun W, Wang Z, et al. FTO suppresses glycolysis and growth of papillary thyroid cancer via decreasing stability of APOE mRNA in an N6-methyladenosine-dependent manner. J Exp Clin Cancer Res. 2022;41(1):42. Published 2022 Jan 28. doi:10.1186/s13046-022-02254-z
Zhou T, Chen G, Chen M, et al. Direct Full-Length RNA Sequencing Reveals an Important Role of Epigenetics During Sexual Reversal in Chinese Soft-Shelled Turtle. Front Cell Dev Biol. 2022;10:876045. Published 2022 Mar 25. doi:10.3389/fcell.2022.876045
Ji X, Sun W, Lv C, et al. ALKBH5-induced circRNA NRIP1 promotes glycolysis in thyroid cancer cells by targeting the miR-541-5p/PKM2 and miR-3064-5p/PKM2 axes. Preprint. Res Sq. 2022;rs.3.rs-1493404. Published 2022 Apr 4. doi:10.21203/rs.3.rs-1493404/v1
Chen YH, Jiang T, Fan BY, et al. Transcriptome-wide N6-methyladenosine methylome profiling of Bombyx mori reveals a potential mechanism of epigenetic regulation on diapause. Preprint. Res Sq. 2022;rs.3.rs-1561012. Published 2022 Apr 18. doi:10.21203/rs.3.rs-1561012/v1
Xu C, Huang H, Zhang M, et al. Methyltransferase-Like 3 Rescues the Amyloid-beta protein-Induced Reduction of Activity-Regulated Cytoskeleton Associated Protein Expression via YTHDF1-Dependent N6-Methyladenosine Modification. Front Aging Neurosci. 2022;14:890134. Published 2022 Apr 25. doi:10.3389/fnagi.2022.890134
Wang X, Lu X, Wang P, et al. SRSF9 promotes colorectal cancer progression via stabilizing DSN1 mRNA in an m6A-related manner. J Transl Med. 2022;20(1):198. Published 2022 May 4. doi:10.1186/s12967-022-03399-3
Zhao TV, Hu Z, Ohtsuki S, et al. Hyperactivity of the CD155 immune checkpoint suppresses anti-viral immunity in patients with coronary artery disease. Nat Cardiovasc Res. 2022;1:634–648. Published 2022 Jul 13. doi:10.1038/s44161-022-00096-8
Li W, Xing C, Bao L, et al. Comprehensive analysis of RNA m6A methylation in pressure overload-induced cardiac hypertrophy. BMC Genomics. 2022;23(1):576. Published 2022 Aug 11. doi:10.1186/s12864-022-08833-w
Chen D, Fu L, Su T, et al. N6-methyladenosine methylation analysis reveals transcriptome-wide expression response to salt stress in rice roots. Environ Exp Bot. 2022;201:104945. Published 2022 Sept. doi:10.1016/j.envexpbot.2022.104945





 Cart (0)
Cart (0)